
The morning was so sparkling that I stopped for a photo opportunity while loading the car. It was -3 according to the new thermometer – it almost never gets this cold. In the greenhouse the poblano/ancho chillies are still producing. The lambs seemed happy enough – after weeks of rain and wind sunny and cold is probably preferable.
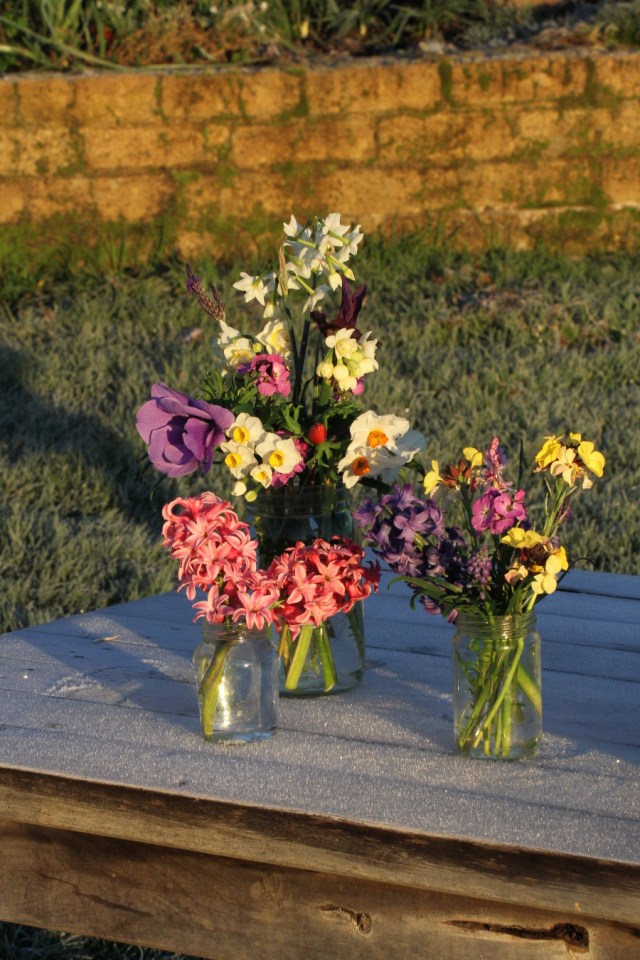
There were a lot of seasonal firsts this week – the first Anemone, the first Dutch Iris, Eye of the Tiger, and the first of my new scented Narcissi, Avalanche and Geranium.

I have always loved the smell of Paper Whites but Geranium (above) is sweeter and more complex.

Avalanche (above) is a sweet little lemon cup.
The shrubby perennial wallflowers, Erysimum, are starting their long bloom period. The lemon one flowers for around 9 months. The big hyacinths are lasting so long in their pots, and smell so sweet in the garden, but several had broken off in the wind so I salvaged them for the vase.

It’s also the first time I have grown Wurmbea stricta or Onixotis Triquetia as it was sold to me (the little spike of pink and magenta above). A South African plant, it has settled in well and has been flowering for a few weeks now.

I planted quite a few new Dutch Iris this year which are all lovely masses of silvery leaves right now, but the first to flower is an older one, I think it is Eye of the Tiger.
Over at Rambling in the Garden, it’s anything but frosty with the hot colours of mid summer flowers.

Ah! Spring!
Love that Iris.
🙂 I really love them, just the leaves alone are gorgeous.
So nice and a cute thought to display them on the frosty table 🙂 Keep warm and don’t leave them out for too long!
Thanks 🙂
The thrill of those first spring flowers is contagious. The Anemone is lovely.
It is lovely, they seem so delicate and temporary.
It is such a pleasant surprise when blog hopping to come to a spring bouquet when we are still sweltering in summer heat going into fall. As our days are shortening yours is lengthening. These posies make me think of what I should be planting this fall for next spring.
Yes it’s good that way. I’m just looking at planting Larkspurs like the ones in Cathy’s vase this week.
Thnaks for sharing the different early narcissi with us – good to see the differences. And the apricot hyacinth is something beacme more appealing the longer I looked at as it’s not a colour I would have considered before – perhaps I will add it to my bulb order… So refreshing to seee your cool colours (without the negative temperatures!) 🙂
I should have a few more daffodils coming on, I’m especially interested to see what the scent of ‘Fragrant Rose’ is like.
Oh, do let us know – sounds too good to be true with a name like that!
Lovely flowers, so nice to get a cool breeze from your garden.
Thank you, it must be really warm where you are now.
The heat index has been 108, yes. very warm.
That’s a chilly morning for you! Brrr!
Quite a few of your blooms have caught my eye: N.’Avalanche’ (I have ‘Geranium’ and adore its scent), I.’Eye of the Tiger’ – gorgeous color! And Onixotis Triquetia is delicate and lovely!
Avalanche is gorgeous with more of the traditional jonquil scent. The onixotis is very delicate and not realising this I have probably doomed it to being overwhelmed by a nearby penstemon. I think I need to move it and add some more.
I’m surprised that the bearded Iris is blooming when it’s still so cold there! It and all your spring blooms are beautiful. I’ve never planted the yellow-flowered Erysimum but will have to try it.
The weather has been so on and off, warm and then cold – the flowers seem to take it in stride.
Beautiful blooms. I can imagine the gorgeous scent too.
Thank you, there are a lot of scents in this vase and to me the ones which are the nicest inside are the daffodil ‘Geranium’ and the yellow Erysimum. It has a nicer scent than the Bowles Mauve.
Beautiful spring flowers and spring sunshine! The onixotis is very pretty. 🙂
All those pretty flowers sure don’t jive with -3 degrees… Yikes! Hope things normalize for you soon, and happy Spring, Cath!
Thanks for the Spring greetings! I’m in degrees C, so -3 while exceptionally cold for us may not be as cold as you are thinking if you are thing in degrees F, which is according to Google 26.6.
Oh no, I’m a Swede, and is far less confused by Celsius than I am by Fahrenheit – I knew what you meant. Either way, it just didn’t seem like a temperature that jived with the kind of flowers you have in your garden. That was the puzzling part for me, but thanks anyway for the clarification! 🙂
So delightful to see your Spring blooms. I don’t want to think about your chilly temperatures I am hoping our Summer will last forever!